Utfordringer til Evolusjonsteorien: Del 2
Del 1 finnes her.
Hva med ikke-darwinistiske prosesser med en utvidet evolusjonær syntese?
Av Günter Bechly, oversatt -herfra ,-ca fra midten.
Bilde 1. Om ikke evolusjon har noe mål, så har evolusjonisme det
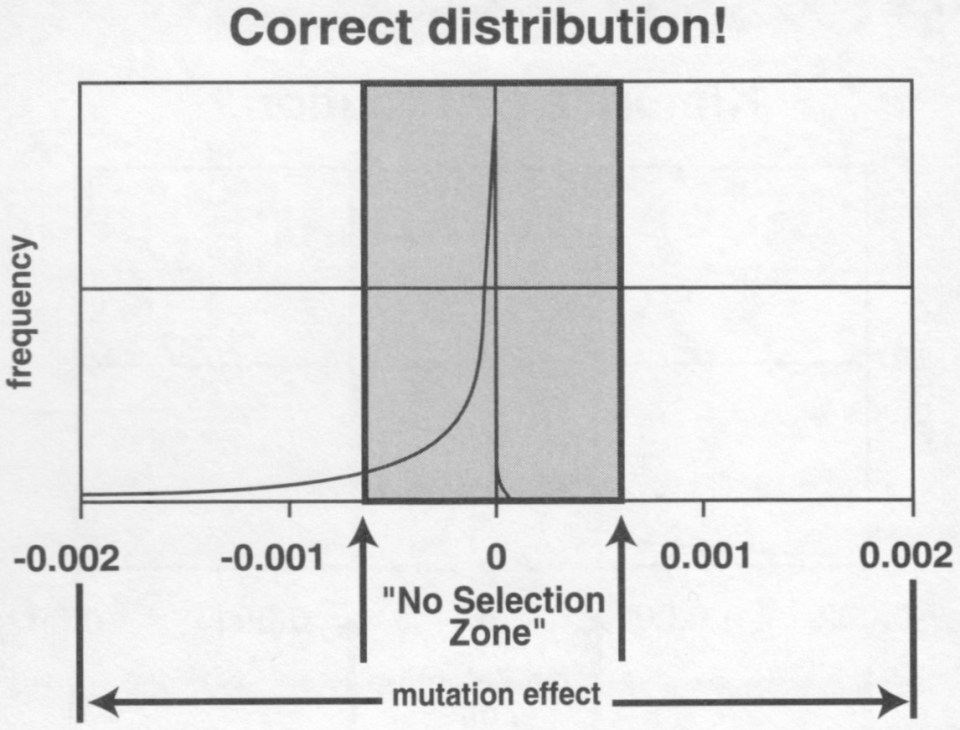
I motsetning til påstandene fra noen forskere om at kritikk av neo-darwinismen er irrelvant fordi neo-darwinisme effektivt er død siden 1960-tallet og vi flyttet utover det for lenge siden (Gould 1980, Swamidass 2018). Neo-darwinisme er ennå langt fra foreldet og representerer det regjerende paradigmet i hovedstrøms evolusjonsbiologi (Hancock et al. 2021). Hancock et al. (2021) konkluderte med at "Den moderne syntesen (eller 'neo-darwinisme'),.., fortsatt er grunnlaget for evolusjonsteori. Neo-darwinisme er i live og og i form."
Likevel, de siste tiårene, innså evolusjonsbiologer mer og mer at den neo-darwinske mekanismen for naturlig/seksuell seleksjon som virker på tilfeldige mutasjoner mislyktes som tilstrekkelig forklaring på opprinnelse til biologisk kompleksitet og mangfold. På konferansen om nye trender innen evolusjonsbiologi som ble holdt i november 2016 på det prestisjetunge Royal Society i London, var hovedinnlegget av østerriksk evolusjonær biolog Gerd Müller, som listet opp fem underskudds-forklaringer til den moderne syntesen (aka neo-darwinisme) inkludert fenotypisk kompleksitet, biologiske nyskapninger, og ikke-graduelle former for overgang (Müller 2016, Bechly et al. 2019). Dermed blir de mest avgjørende fenomenene i makroevolusjon uten tvil ikke forklart av neo-darwinisme.
På samme måte etterlot kunngjøringen fra konferansen 2018 i Salzburg av New York Academy of Sciences, ikke rom for tvil: "I mer enn et halvt århundre har det blitt akseptert at ny genetisk informasjon for det meste er avledet fra tilfeldig‚ feilbaserte begivenheter. Nå er det anerkjent at feil ikke kan forklare genetisk nyhet og kompleksitet "(Witzany 2018, 2020). Denne erkjennelsen var gnisten til kunngjøringen av en tredje evolusjonsvei(Shapiro et al. 2014) og en utvidet evolusjonær syntese (f.eks. Laland et al. 2015, Garte 2016, Müller 2017), som anerkjente de forklarende underskuddene i den  moderne syntesen og antydet flere andre evolusjonsmekanismer som fenotypisk plastisitet (evolvabilitet), nisjekonstruksjon, epigenetikk, naturlig genetisk ingeniørvitenskap, hybridogenese, symbiogenese og neo lamarckisme. I motsetning til feiende påstander (f.eks. Müller 2013), forklarer ingen av disse mekanismene opprinnelsen til ny spesifisert informasjon og komplekse tilpasninger noe bedre enn neo-darwinisme gjorde, eller rettere ikke sagt klarte å gjøre. Ingen av disse alternative mekanismene løser eller endog adresserer ventetidsproblemet.
moderne syntesen og antydet flere andre evolusjonsmekanismer som fenotypisk plastisitet (evolvabilitet), nisjekonstruksjon, epigenetikk, naturlig genetisk ingeniørvitenskap, hybridogenese, symbiogenese og neo lamarckisme. I motsetning til feiende påstander (f.eks. Müller 2013), forklarer ingen av disse mekanismene opprinnelsen til ny spesifisert informasjon og komplekse tilpasninger noe bedre enn neo-darwinisme gjorde, eller rettere ikke sagt klarte å gjøre. Ingen av disse alternative mekanismene løser eller endog adresserer ventetidsproblemet.
En annen ikke-darwinistisk eller rettere sagt ikke-adaptasjonistisk tilnærming ble antydet av talsmenn for nøytral evolusjon (Kimura 1968, 1983, Jensen et al. 2019), som la vekt på rollen som vektla genetisk drift snarere enn seleksjon. I motsetning til de dristige påstandene fra noen forskere (Duret 2008, Koonin 2016, Moran 2018, Swamidass 2018), har nøytral teori i det hele tatt ikke blitt det nye hovedstrøms-synet i evolusjonsbiologien, men har heller vist seg å være helt irrelvant for makroevolusjon. Dette er neppe overraskende ettersom nøytral teori hovedsakelig gjelder molekylær evolusjon. I en fersk anmeldelse kom Kern & Hahn (2018) til en sviende-konklusjon:
Bilde 2. Vitenskapsfilosof etterlyser bevis på at mekroevolusjon har forekommet
"I dette perspektivet evaluerer vi den forklarende kraften til den nøytrale teorien om molekylær evolusjon, 50 år etter introduksjonen av Kimura. Vi argumenterer for at den nøytrale teorien ble støttet av upålitelige teoretiske og empiriske bevis fra begynnelsen, og at vi i lys av moderne, genomskala-data kan avvise dens universalitet. ... På 50 -årsjubileet for den nøytrale teorien om molekylær evolusjon, har vi blitt utfordret til oppgaven med å spørre: hvordan har den nøytrale teorien bleknet i lys av adaptiv variasjon i og mellom arter? Med et ord, dårlig. … Den nøytrale teoriens forklarende kraft har aldri vært eksepsjonell. … Hver av de opprinnelige bevislinjene for den nøytrale teorien er nå forfalsket, ... som en konsekvens, tror vi at den nøytrale teorien overveldende er avvist ".
 Selv om nøytrale prosesser som genetisk drift kan forklare noen former for spesiasjon ved akkumulering av forskjeller i separerte linjer, kan de absolutt ikke forklare integrert og spesifisert adaptiv biologisk kompleksitet.
Selv om nøytrale prosesser som genetisk drift kan forklare noen former for spesiasjon ved akkumulering av forskjeller i separerte linjer, kan de absolutt ikke forklare integrert og spesifisert adaptiv biologisk kompleksitet.
Derfor ble en ny tilnærming kalt 'konstruktiv nøytral evolusjon' (KNE) foreslått som en skralle med den påståtte kreative kraften til å få til ny biologisk (spesielt cellulær) kompleksitet og til og med tilsynelatende irreduserbar kompleksitet (Stoltzfus 1999, 2012, Gray et al. 2010, Lukeš et al. Imidlertid feilet de, ikke bare ved ikke å bli omfavnet med stor entusiasme av evolusjonsbiologer (Muñoz-Gómez et al. 2021), men representerer heller en ignorert frynseteori, sannsynligvis fordi den bare kan forklare Rube-Goldberg-maskiner, ineffektive og unødig komplekse, (kanskje som blodproppkaskaden), men ikke svært tilpasningsdyktig og effektive komplekse nye organer (for eks. motstrømvarme-utvekslingssystemet i hvaler) og molekylære maskiner som bakterie flagellen. Faktisk kan KNE best øke kompleksiteten og gjensidig avhengighet, men ikke tilpasning og funksjonalitet, slik at den må forutsette allerede eksisterende funksjon før en ytterligere akkumulering av kompleksitet (Jones 2011).
Bilde 3. Kimura var talsmann for nøytral evolusjon
Speijer (2011) konkluderte med at CNE "har store konseptuelle problemer, [og] kan ikke forklare observerte mønstre av komplekse prosesser". Når det gjelder ALGE-tilfeller av cellularkompleksitet generert av KNE, innrømmet selv den siste gjennomgangen (Muñoz-Gómez et al. 2021) at "de fleste, om ikke alle disse foreslåtte eksemplene forblir spekulative og venter på ytterligere bevis og detaljerte evolusjonsfortellinger". Problemet blir til og med verre når koordinerte mutasjoner vil bli krevd, som Pillai et al. (2020) fant at "i prinsippet kunne molekylære samlinger oppstå og bli mer komplekse via nøytrale prosesser, men dette scenariet er usannsynlig hvis det er nødvendig med mange mutasjoner". Derfor unnlater KNE også å gi noen potensiell løsning på ventetidsproblemet. Tvert imot, nøytrale prosesser ville kraftig blåse opp fikseringstidene som ble demonstrert av Hössjer et al. (2021), som fant at "den forventede ventetiden øker eksponentielt med m, for en nøytral seleksjons-modell, når tilbake-mutasjoner er mulig".
 Noen forskere mener feilaktig at teorien om punktert likevekt har forklart plutselig opprinnelse til biologisk nyheter, og derfor kan løse ventetidsproblemet. Ingenting kunne vært lenger fra sannheten. Punktert likevekt ble introdusert av Eldredge & Gould (1972) som en forklaring på Stasis (Gould 1991) og diskontinuiteter i fossilprotokollen til (mikroevolusjonære) art-til-artsovergang og komplekse biologiske nyheter. Den vanlige misforståelsen av punktert likevekt som en saltasjonistisk, makroevolusjonær teori, snarere enn en gradvis mikroevolusjonær teori, kan ha blitt provosert av en villedende originaltittel: Punktert likevekt: Et alternativ til fyletisk gradualisme av Eldredge og Gould (1972) for deres originalartikkel. . Løvtrup (1981) hadde allerede avvist punktert likevekt, nettopp fordi det fremdeles var en form for gradvishet og ikke en makro-mutasjonsteori favorisert av ham. Han konkluderte med at "Eldredge og Goulds teori har ikke noe med makroevolusjon å gjøre i Goldschmidts forstand". Selvfølgelig sympatiserte Stephen Jay Gould (1977, 1980) faktisk med saltasjonsideer og tenkte at "makroevolusjon gikk videre ved den sjeldne suksessen til disse håpefulle monstrene, ikke av kontinuerlige små endringer i populasjonen", men dette var uavhengig av og helt uten tilknytning til hans hans egen 'punk eek' teori (Moran 2008). Gould (1982) la eksplisitt vekt på at "punktert likevekt ikke er en teori om makromutasjon", som burde ha avsluttet problemet. Likevel ble Ernst Mayr (1982: 617) forvirret og hevdet feilaktig at Gould og Eldredges teori om punktert likevekt innebærer saltasjoner, som tilsvarer Goldschmidts 'Håpefulle monstre'.
Noen forskere mener feilaktig at teorien om punktert likevekt har forklart plutselig opprinnelse til biologisk nyheter, og derfor kan løse ventetidsproblemet. Ingenting kunne vært lenger fra sannheten. Punktert likevekt ble introdusert av Eldredge & Gould (1972) som en forklaring på Stasis (Gould 1991) og diskontinuiteter i fossilprotokollen til (mikroevolusjonære) art-til-artsovergang og komplekse biologiske nyheter. Den vanlige misforståelsen av punktert likevekt som en saltasjonistisk, makroevolusjonær teori, snarere enn en gradvis mikroevolusjonær teori, kan ha blitt provosert av en villedende originaltittel: Punktert likevekt: Et alternativ til fyletisk gradualisme av Eldredge og Gould (1972) for deres originalartikkel. . Løvtrup (1981) hadde allerede avvist punktert likevekt, nettopp fordi det fremdeles var en form for gradvishet og ikke en makro-mutasjonsteori favorisert av ham. Han konkluderte med at "Eldredge og Goulds teori har ikke noe med makroevolusjon å gjøre i Goldschmidts forstand". Selvfølgelig sympatiserte Stephen Jay Gould (1977, 1980) faktisk med saltasjonsideer og tenkte at "makroevolusjon gikk videre ved den sjeldne suksessen til disse håpefulle monstrene, ikke av kontinuerlige små endringer i populasjonen", men dette var uavhengig av og helt uten tilknytning til hans hans egen 'punk eek' teori (Moran 2008). Gould (1982) la eksplisitt vekt på at "punktert likevekt ikke er en teori om makromutasjon", som burde ha avsluttet problemet. Likevel ble Ernst Mayr (1982: 617) forvirret og hevdet feilaktig at Gould og Eldredges teori om punktert likevekt innebærer saltasjoner, som tilsvarer Goldschmidts 'Håpefulle monstre'.
Bilde 4. Ulike linjer trekkes ut fra ulike hypoteser
 Denne grove misforståelsen ble opprinnelig adoptert og foreviget av Daniel Dennett, som først senere erkjente og korrigerte sin feil (Dennett 1995). Et salasjonistisk syn på punktert likevekt ble eksplisitt avvist av Gould & Eldredge (1986), som avklarte at "vår største frustrasjon gjennom denne debatten - var den vedvarende forvirringen, spesielt i pressen, av punktert likevekt med ekte saltasjonisme". Gould (1991) sa at "enkle misforståelser om grunnleggende innhold var urovekkende vanlig, selv blant profesjonelle evolusjonister. Mange kolleger trodde at vi hadde dratt fram det gamle anti-darwinsistiske spekteret av makromutasjonisme, eller virkelig plutselig spesifikasjon i en enkelt generasjon av en stor og utrolig heldig mutasjon. Jeg vet ikke hvorfor dette skjedde ". På samme måte beklaget Eldredge (2008) over at "artikkelen vakte en bølge av oppmerksomhet - mye av det negativt, som ble beskyldt for å dreie mot Darwin og for å fremme en form for 'saltasjonisme' - diskrediterte ideer om evolusjonsprosess i plutselige 'hopp' ('Saltus' er det latinske ordet for 'hopp') gjennom en ukjent genetisk mekanisme eller annet '. Bortsett fra det, klarte ikke punktert likevekt å overbevise mange evolusjonsbiologer om å være en nyttig idé i det hele tatt. Dawkins (1986) kalt punktert likevekt en "mindre rynke på overflaten av neo-darwinsk teori". Hancock et al. (2021) konkluderte med at "punktert likevekt ikke representerte en stor revolusjon innen evolusjonsbiologi", og Coyne (2021) erklærte til og med punktert likevekt for å være en død teori. Uansett hva, selv om punktert likevekt ville være riktig og gjelde for makroevolusjon, vil det heller gjøre ventetidsproblemet verre, fordi det ville antyde overganger som var så raske at de unngikk fossilprotokollen og de nødvendige genetiske endringene fra å stamme fra og spre seg i en forfedres befolkning. Dermed er punktert likevekt ikke en løsning, men snarere bare et synonym for ventetidsproblemet.
Denne grove misforståelsen ble opprinnelig adoptert og foreviget av Daniel Dennett, som først senere erkjente og korrigerte sin feil (Dennett 1995). Et salasjonistisk syn på punktert likevekt ble eksplisitt avvist av Gould & Eldredge (1986), som avklarte at "vår største frustrasjon gjennom denne debatten - var den vedvarende forvirringen, spesielt i pressen, av punktert likevekt med ekte saltasjonisme". Gould (1991) sa at "enkle misforståelser om grunnleggende innhold var urovekkende vanlig, selv blant profesjonelle evolusjonister. Mange kolleger trodde at vi hadde dratt fram det gamle anti-darwinsistiske spekteret av makromutasjonisme, eller virkelig plutselig spesifikasjon i en enkelt generasjon av en stor og utrolig heldig mutasjon. Jeg vet ikke hvorfor dette skjedde ". På samme måte beklaget Eldredge (2008) over at "artikkelen vakte en bølge av oppmerksomhet - mye av det negativt, som ble beskyldt for å dreie mot Darwin og for å fremme en form for 'saltasjonisme' - diskrediterte ideer om evolusjonsprosess i plutselige 'hopp' ('Saltus' er det latinske ordet for 'hopp') gjennom en ukjent genetisk mekanisme eller annet '. Bortsett fra det, klarte ikke punktert likevekt å overbevise mange evolusjonsbiologer om å være en nyttig idé i det hele tatt. Dawkins (1986) kalt punktert likevekt en "mindre rynke på overflaten av neo-darwinsk teori". Hancock et al. (2021) konkluderte med at "punktert likevekt ikke representerte en stor revolusjon innen evolusjonsbiologi", og Coyne (2021) erklærte til og med punktert likevekt for å være en død teori. Uansett hva, selv om punktert likevekt ville være riktig og gjelde for makroevolusjon, vil det heller gjøre ventetidsproblemet verre, fordi det ville antyde overganger som var så raske at de unngikk fossilprotokollen og de nødvendige genetiske endringene fra å stamme fra og spre seg i en forfedres befolkning. Dermed er punktert likevekt ikke en løsning, men snarere bare et synonym for ventetidsproblemet.
Bilde 5. En propaganda-teknikk som stadig benyttes
Til slutt erkjente noen forskere (Davidson & Erwin 2006), at mikroevolusjonære prosesser ikke kan utgjøre store forskjeller i kroppsplaner, i stedet antydet de at små og enkle endringer i regulatoriske kontrollgener kunne produsere store fenotypiske endringer av kroppsplanene, uten at noen nye gener eller proteiner utviklet seg. Derfor hevdet Marshall (2013) at "ny phyla ikke ble laget av nye gener, men i stor grad dukket opp gjennom restrukturering av genreguleringsnettverkene (GRN) av allerede eksisterende gener". Et lignende syn ble foreslått av Cabej (2019) som løsning på problemet med den brå opprinnelsen til nye dyrekroppsplaner i den kambriske eksplosjonen. Imidlertid har denne muligheten ikke bare blitt kritisert av andre evolusjonsbiologer (f.eks. Alonso og Wilkins 2005, Hoekstra og Coyne 2007, Stern & Orgogozo 2008), men har faktisk blitt empirisk tilbakevist av nyere fylogenomiske studier (f.eks. & Holland 2018, Hegerge 2018, Hegerge 2018, Hegerge 2018, Hegery 2018, Hegery 2018, Hegery 2018, Hegery 2018, Hegery 2018, Hegery 2018, Heger. et al. Paps & Holland (2018) konkluderte med at "i motsetning til det rådende synet, avdekker dette en enestående økning i omfanget av genomiske nyheter, i opprinnelsen av metazoaner, og identifiserer 25 grupper av metazoanspesifikke gener som er essensielle på tvers av dyreriket".
 Paps (2018) uttrykte det enda tydeligere at "rekonstruering av genomet til den siste felles stamfaren til eksisterende dyr har avduket en enestående fremvekst av nye gener, og fremhever rollen til genomisk nyheter i opprinnelsen til metazoans". Tilsvarende har Heger et al. (2020) "identifisert 157 bilaterisk-spesifikke gener", som "motsier det nåværende synet [og] avslører at gener med bilaterisk opprinnelse er robust assosiert med nøkkelfunksjoner i eksisterende bilaterianer". Det er klart at nye komplekse organer og vev krevde nye gener og nye proteiner, og ikke bare omfordeling av eksisterende genetiske 'Lego-klosser'. Så ventetidsproblemet kan ikke forklares bort som et pseudo -problem. Bortsett fra det, vil vi vise i vår kommende studie (Bechly et al. i prep.) At til og med den cis-regulerende hypotesen innebærer et dyptgående ventetidsproblem. Opprinnelsen og spredningen av genetiske innovasjoner som korrelerte med makroevolusjonære overganger krever en forklaring innenfor begrensningene, gitt av populasjons-genetikk og fossilregisteret.
Paps (2018) uttrykte det enda tydeligere at "rekonstruering av genomet til den siste felles stamfaren til eksisterende dyr har avduket en enestående fremvekst av nye gener, og fremhever rollen til genomisk nyheter i opprinnelsen til metazoans". Tilsvarende har Heger et al. (2020) "identifisert 157 bilaterisk-spesifikke gener", som "motsier det nåværende synet [og] avslører at gener med bilaterisk opprinnelse er robust assosiert med nøkkelfunksjoner i eksisterende bilaterianer". Det er klart at nye komplekse organer og vev krevde nye gener og nye proteiner, og ikke bare omfordeling av eksisterende genetiske 'Lego-klosser'. Så ventetidsproblemet kan ikke forklares bort som et pseudo -problem. Bortsett fra det, vil vi vise i vår kommende studie (Bechly et al. i prep.) At til og med den cis-regulerende hypotesen innebærer et dyptgående ventetidsproblem. Opprinnelsen og spredningen av genetiske innovasjoner som korrelerte med makroevolusjonære overganger krever en forklaring innenfor begrensningene, gitt av populasjons-genetikk og fossilregisteret.
Bilde 6. Det kreves et visst minimums-sett av gener for livet
Vi kan trygt konkludere med at ingen av de nylig foreslåtte mekanismene adresserer og løser opprinnelsen til komplekse nye organer og det underforståtte ventetidsproblemet for de genetiske endringene som kreves for å oppnå en slik ombygging av kroppsplanen. Derfor er det teoretiske tomrommet etter at neo-darwinismens mislyktes fortsatt ikke fylt ut og krever en større omtenking av teorien (Laland et al. 2014). Nye paradigmer bør vurderes og utforskes som for eksempel kan omfatte adaptive makromutasjoner som ikke er tilfeldige, som antydet av talsmenn for kvanteutvikling (McFadden & Al-Khalili 1999, McFadden 2001, og Ryzko 2009, Elsheik 2016, versjoner av saltasjon og mutasjon i ER. ånden i Goldschmidts 'Håpefulle monstre' -hypotese (Gould 1977, Løvtrup 1987, Theißen 2006, Rieppel 2001, 2017). I denne sammenhengen er det interessant at det rulende paradigmet for tilfeldige mutasjoner blir mer og mer utfordret og tilbakevist av eksperimentelle bevis (Sniegowski 1995, Wright 2000, Jablonka & Lamb 2005, Kokić 2010, Merlin 2010, Shapiro 2011, 2013, Livnat 2013, Martincorena & Luscombe 2013, Fitzgerald & Rosenberg 2019, Monroe et al.
Arts-par utfordringen
 Endelig er det også et annet problem som hittil i stor grad har blitt oversett: Den morfologiske likheten til moderne artspar som har divergert i en sammenlignbar tidsramme, utgjør et halt problem, fordi det innebærer at de makrovolusjonære prosessene som var på jobb og vanlig i historien til livet, i alle perioder av jordhistorien og alle grupper av organismer, tilsynelatende var helt fraværende i opprinnelsen til alle millioner av levende arter. For å drive gjennom dette poenget, undersøkte TimeTree.org -lenke https://timetree.org/ (Hedges og Kumar 2009, Hedges et al. 2006, 2015, Kumar et al. 2017), som er en databank av 148.876 levende arter av alle slag eller organismer, med molekylære klokkeestimater av deres tid for divergens, basert på 4.185 studier. Når du sonderer noen par av arter, selv de med lengre divergenstider enn tilgjengelig, for utvikling av kroppsplanforskjeller mellom f. eks. pakiketer og basilosaurider (atskilt med omtrent 4-5 millioner år), finner vi uten unntak at morfologiene deres neppe kan adskilles for lekfolk, og de kan ofte fortsatt hybridisere.
Endelig er det også et annet problem som hittil i stor grad har blitt oversett: Den morfologiske likheten til moderne artspar som har divergert i en sammenlignbar tidsramme, utgjør et halt problem, fordi det innebærer at de makrovolusjonære prosessene som var på jobb og vanlig i historien til livet, i alle perioder av jordhistorien og alle grupper av organismer, tilsynelatende var helt fraværende i opprinnelsen til alle millioner av levende arter. For å drive gjennom dette poenget, undersøkte TimeTree.org -lenke https://timetree.org/ (Hedges og Kumar 2009, Hedges et al. 2006, 2015, Kumar et al. 2017), som er en databank av 148.876 levende arter av alle slag eller organismer, med molekylære klokkeestimater av deres tid for divergens, basert på 4.185 studier. Når du sonderer noen par av arter, selv de med lengre divergenstider enn tilgjengelig, for utvikling av kroppsplanforskjeller mellom f. eks. pakiketer og basilosaurider (atskilt med omtrent 4-5 millioner år), finner vi uten unntak at morfologiene deres neppe kan adskilles for lekfolk, og de kan ofte fortsatt hybridisere.
Her er noen eksempler fra TimeTree (dateringen er oppdatert sammenlignet med Bechly 2022A, basert på den nyeste versjonen av TidsTre-databasen):
*Gran (Abies Spec.) Og edelgran (Cedrus Spec.) Tilhører samme underfamilie av bartrær, men skilte seg allerede for 136 millioner år siden;
*Vanlig huflue (Musca Domestica) og liten husflue (Fannia Scalaris) divergerte 39-45 for mill. år siden;
*Northern Damsel-flue (Coenagrion Hastulatum) og Azure Damsel-flue (Coenagrion Pauella) divergerte for 11.8 mill. år siden;
*Vanlig europeisk frosk (Rana Temporaria) og Moor Frog (Rana Arvalsis) divergerte for 13.2 mill. år siden;
Bilde 7. Det er grenser for hvor langt unna en art beveger seg på 5 mill. år
*Galapagos land iguana (Conolophus spec.) Og Galapagos marine iguanaer (Amblyrhynchus spec.) Divergerte for 23.9-24.2 mill. år siden (Marine iguanaer kan utskille salt fra en kjertel ved neseborene og har en flatere hale, men ser ellers ut veldig lik kusinen sin.);
*Grønnsanger (Phylloscopus nitidus) og Bonellisanger (Phylloscopus Bonelli) divergerte for 15,2 mill. år siden (minst 4-7 millioner år ifølge Helbig et al. 1995), men ser nesten identiske ut og kan fremdeles hybridisere (denne mønster er typisk for fugler);
* Gråspurv (passer Domesticus) og trespurv (passer Montanus) divergerte for 5.6 mill. år siden og kan fremdeles hybridisere i naturen;
*Husmus (Mus Musculus) og rotte (Rattus norvegicus) divergerte 11,6-13,1 mill. år siden (minst 12 millioner år ifølge Kimura et al. 2015);

*Europeisk bison (Bison Bonasus) og huskveg (Bos Taurus) divergerte for 3,56 mill. år siden og kan fremdeles hybridisere som beefalos (US)-Bilde 8: beefalo.
*Hester (Equus Caballus) og esler (Equus Asinus) divergerte for 11.1 mill. år siden og kan fremdeles hybridisere som muldyr;
*Asiatisk elefant (Elephas Maximus) og afrikansk elefant (Loxodonta Africana) divergerte for 8,4 mill. år siden (minst 7,6 millioner år ifølge Rohland et al. 2007), og til og med afrikanske Savanne -elefanter (L. Africana) og den veldig like skogselefanten (L.Cyclotis) divergerte for 5,5 mill. år siden (minst 4 millioner år ifølge Rohland et al. 2007);
*Brillebjørner (Tremarctos Ornatus) og Asiatiske svartbjørner (Ursus / Selenarctos Thibetanus) divergerte for 13,9 millioner år siden og kan fortsatt hybridisere i fangenskap (Mondolfi & bodde 1981);
*ELveoter (Lutra Lutra) og brunsel (Arctocephalus Pusillus) divergerte for 40 mill. år siden (og er fremdeles langt fra forskjellen mellom pakiketer og basilosaurider, som skulle ha foregått på en tidel av tiden);
*Elve-flodhest (Hippopotamus Amphibius) og pygme-flodhest (Choeropsis Liberiensis) divergerte for 7 mill. år siden (flodhester representerer de nærmeste levende pårørende til hvaler);
*Vanlig delfin (Delphinus Delphis) og Flaske Nese Delfin(Tursiops truncatus) divergerte 2,25 mill. år siden (dette er mer enn det dobbelte av den tilgjengelige tiden for transformasjon av firbente protoksider til fullt marine pelagicetider, men har bare oppnådd mindre forskjeller).
 De fleste av disse nylige artsparene er bare forskjellige i allometriske forstand samt mindre fargemønstre. De ser vanligvis så like ut at de knapt kunne bli atskilt av lekfolk, selv om de var adskilt i mye lengre tid enn det som var tilgjengelig for de fleste store overganger i fossilregisteret.
De fleste av disse nylige artsparene er bare forskjellige i allometriske forstand samt mindre fargemønstre. De ser vanligvis så like ut at de knapt kunne bli atskilt av lekfolk, selv om de var adskilt i mye lengre tid enn det som var tilgjengelig for de fleste store overganger i fossilregisteret.
Så hva med store aper og mennesker? Sjimpanse (Pan Paniscus) og Gorilla (Gorilla Gorilla) divergerte ifølge TimeTree for 8,6 millioner år siden og mennesker (Homo Sapiens) fra sjimpanser for 6,4 millioner år siden, noe stemmer overens med Hominin fossilregisteret. Det er to muligheter: Enten følger du forskerne som anser den biologiske forskjellen mellom mennesker og sjimpanser som marginale. Da vil dette eksemplet bare bekrefte mønsteret beskrevet ovenfor. Eller du anser mennesker som veldig forskjellige fra sjimpanser, basert på deres forskjellige bipedale bevegelse og spesielt deres mentale kapasitet og kulturelle prestasjoner. I det siste tilfellet ville mennesker representere det eneste unntaket fra mønsteret som vi kunne finne, som uten tvil vil representere en bemerkelsesverdig bekreftelse av jødisk-kristen, menneskelig eksepsjonalisme.
Bilde 9. Store forskjeller -når det kommer til kromosomer
Disse eksemplene kan utvides uendelig, men bør være tilstrekkelig for å fastsette poenget. Det er helt klart grenser for hva ikke-styrt evolusjon kan gjøre i løpet av noen få millioner år, og disse grensene er langt under nivået på noen store kroppsplanoverganger. Dermed kan vi trygt konkludere med at det er to udiskutable fakta som krever en tilstrekkelig forklaring:
1.) Det er mange eksempler på fossile artspar med veldig forskjellige kroppsplaner som divergerte innenfor et tid på 5 (± 5) millioner år. Dette er enda mer bemerkelsesverdig hvis vi vurderer at det bare er rundt 350 000 beskrevne fossile arter (ekstrapolert basert på data i Teichert 1956, Valentine 1970, Raup 1976 og Alroy 2002), som bare representerer en liten brøkdel av de estimerte 5-50 milliarder arter som noen gang har bodd på jorden (Raup 1991).
2.) Det eksisterer ingen levende artspar med selv fjernt lignende forskjeller i kroppsplan som er datert for å ha avviket i en lignende tidsramme. Dette er enda mer bemerkelsesverdig hvis vi vurderer at det er anslagsvis 8,7 millioner levende arter (Mora et al. 2011, Strain 2011, Sweetlove 2011), hvorav mer enn 2 millioner er beskrevet (IISE 2012). Tidligere estimater av det totale antallet levende arter varierte fra 3-100 millioner arter (mai 1988, Tangley 1997, Chapman 2009), men hvis mikrober er inkludert, kan det til og med være opp til en billion levende arter (Locey & Lennon 2016, Latty & Lee 2019).
Tatt i betraktning at tidsvinduet er på bare 5-10 millioner år for de fleste av av de plutselige-oppdukkingenen av nye kroppsplaner i fossilregisteret (Bechly & Meyer 2017, Bechly 2021), den bayesiske sannsynligheten for ikke å finne et eneste eksempel på lignende morfologisk forskjell, som har sin opprinnelse på en lignende tidsramme blant de millioner av levende arter, er i utgangspunktet nær null. Med tanke på dette enkle argumentet som en siste spiker i kisten til darwinistisk ikke-styrt evolusjon.
Bilde 10. Eks. på underliggende filosofis betydning
 Basert på dette argumentet stilte jeg formelt og offentlig følgende utfordring til evolusjonsbiologer (Bechly 2022a): Finn i den enorme databasen med nesten 149 000 arter på Timetree.org bare et enkelt eksempel på et hvilket som helst par forskjellige arter som har divergert for rundt 5 millioner år siden (gi eller ta noen millioner år) i henhold til en enighet om multiple molekylære klokkeundersøkelser, og som viser en morfologisk forskjell i kroppsplanene deres som kan sammenlignes med, for eksempel, Pakicetus og Basilosaurus. For å være tydelig har selvfølgelig ingen evolusjonsbiolog noen gang hevdet at Pakicetus var den faktiske stamfaren til Basilosaurus. Det representerte heller en sidegren av Cetacean stamgruppen. Men det innebærer definitivt at stamarten var omtrent lik i kroppsplan til Raoellidae og Pakictheae. Derfor er denne utfordringen helt gyldig og rimelig.
Basert på dette argumentet stilte jeg formelt og offentlig følgende utfordring til evolusjonsbiologer (Bechly 2022a): Finn i den enorme databasen med nesten 149 000 arter på Timetree.org bare et enkelt eksempel på et hvilket som helst par forskjellige arter som har divergert for rundt 5 millioner år siden (gi eller ta noen millioner år) i henhold til en enighet om multiple molekylære klokkeundersøkelser, og som viser en morfologisk forskjell i kroppsplanene deres som kan sammenlignes med, for eksempel, Pakicetus og Basilosaurus. For å være tydelig har selvfølgelig ingen evolusjonsbiolog noen gang hevdet at Pakicetus var den faktiske stamfaren til Basilosaurus. Det representerte heller en sidegren av Cetacean stamgruppen. Men det innebærer definitivt at stamarten var omtrent lik i kroppsplan til Raoellidae og Pakictheae. Derfor er denne utfordringen helt gyldig og rimelig.
Det er ingen tenkelig grunn til at en forskjell som den mellom Pakicetus og Basilosaurus skal være begrenset til fossilprotokollen, der den kan finnes i en rekke eksempler blant alle grupper av organismer, samtidig som det totalt er fraværende blant millioner av nyere arter. Så utfordringen kan til og med formuleres på en mer avslappet og sjenerøs måte, som ikke er begrenset til TimeTree -databasen. Bare finn et par arter blant de millioner levende artene for å møte utfordringen; Bare et enkelt eksempel! Hvis ikke-styrt evolusjon virkelig kan gjøre sin magi, bør ikke dette være for vanskelig. Imidlertid, hvis utfordringen ikke kan oppfylles, må Darwinister bli bedt om å forklare hvorfor.
En åpenbar mulig innvending mot artsparutfordringen kan være, at slike nylige artspar ikke representerer forfedre-etterkommer avstamning, men bare søster- eller fetterlinjer som begge divergerte fra en felles stamfar. Imidlertid gjelder dette også for de fleste fossile eksempler, og gjør faktisk saken enda mer problematisk. Mens forskjeller i forfedre-etterkommer linjer bare kunne samle seg i en enkelt utviklende avstamning, kunne søsterlinjer begge utvikle seg forskjellig fra samme tid, og burde dermed heller presentere mer og ikke mindre morfologisk ulikhet. Tydeligvis mislyktes faktiske forklaringer i tilfellet med store overganger i fossilregisteret . Dette representerer uten tvil en kraftig uavhengig empirisk bekreftelse for hypotesen om at mikroevolusjonære (populasjonsgenetiske) prosesser ikke kan ekstrapoleres over store perioder, som tilstrekkelig forklaring på opprinnelsen til komplekse biologiske nyheter i livets historie. {sml. bror til Kjell Aukrusts utregning om jomsvikingenes høyde -oversetters tilføyelse.}
 Kanskje vil evolusjonister appellere til ennå ukjente ikke-darwinistiske prosesser (se over). Den store fordelen med dette nye argumentet er imidlertid at det er totalt uavhengig av arten av transformasjonsprosessen. Du kan ganske enkelt betrakte prosessen som en svart boks. Derfor er det helt irrelevant hvis Darwinister oppfinner en ny mulig mekanisme. Det avgjørende poenget er ikke prosessen, men at det resulterende mønsteret av ny kroppsplaner konsekvent har blitt brått til i den fjerne fortiden, men ikke i nyere fortid.
Kanskje vil evolusjonister appellere til ennå ukjente ikke-darwinistiske prosesser (se over). Den store fordelen med dette nye argumentet er imidlertid at det er totalt uavhengig av arten av transformasjonsprosessen. Du kan ganske enkelt betrakte prosessen som en svart boks. Derfor er det helt irrelevant hvis Darwinister oppfinner en ny mulig mekanisme. Det avgjørende poenget er ikke prosessen, men at det resulterende mønsteret av ny kroppsplaner konsekvent har blitt brått til i den fjerne fortiden, men ikke i nyere fortid.
Så hvordan reagerte evolusjonsbiologer på denne nye utfordringen. Stort sett gnisninger, bortsett fra en viss diskusjon på Peaceful Science, der prof. Arthur Hunt, en botaniker og geolog (soil scientist) fra University of Kentucky, antydet at en gruppe hawaiiske endemiske planter kalt Hawaiian Silversword Alliance møter utfordringen. Jeg tilbakeviste denne påstanden i en artikkel (Bechly 2022b), som viste at de helt forskjellige vekstformene for det meste skyldes fenotypisk plastisitet og til og med forekommer innenfor samme art. Igjen ingenting annet enn gnisning. Et år senere står utfordringen fremdeles og mangler alvorlig respons, noe som er ganske illustrerende og viser at det hevede problemet er ekte og et gyldig problem, som krever en adekvat forklaring.
Bilde 11. Darwin ga flere kriterier for krtikk -i forhold til egen teori
Etter mitt syn tyder de kumulative bevisene på at den eneste adekvate forklaring er at darwinisme er feilaktig, og dette gjelder ikke bare den neo-darwinistiske prosessen med tilfeldig mutasjon og naturlig seleksjon, men ethvert syn kalt utvidet syntese (f.eks. Shapiro et al. 2014 , Laland et al. Det er ingen evolusjonær grunn til at den kreative kraften i denne prosessen skulle ha vært aktiv over hele jordhistorien, men deretter sluttet å fungere i løpet av de siste 10 millioner årene. Intelligente design -talsmenn kan lett forklare dette mønsteret: Det fantes kreativ intelligent intervensjon i livets historie, men denne kreative aktiviteten opphørte bevisst med menneskers ankomst som de endelige målskivene. Enhver ytterligere identifisering av den intelligente årsaken vil måtte overtråkke de metodologiske grensene for design-slutningen, men jødisk-kristne teister vil helt sikkert anerkjenne en skremmende korrespondanse med det bibelske budskapet, som sier at Gud hvilte fra sin kreative aktivitet etter skapelsen av mennesker ( (1. Mosebok 2: 2-3).
En ny utfordring: Det kollapsende treproblemet
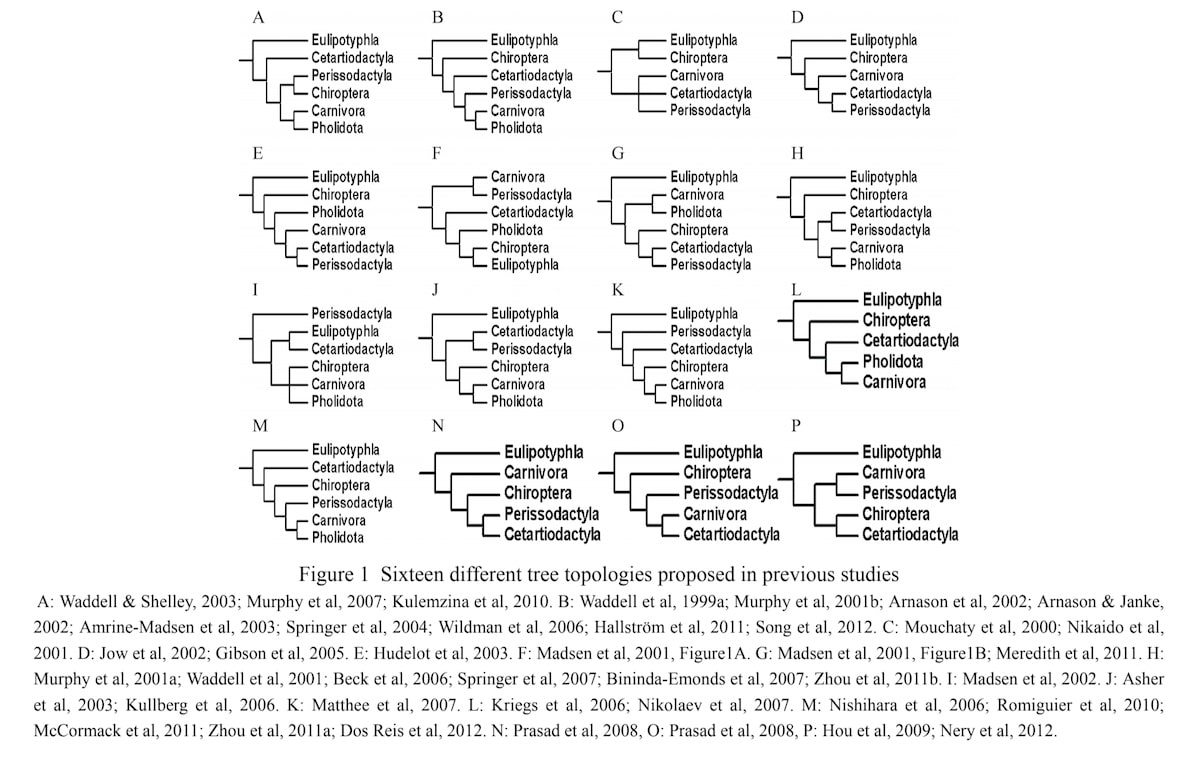 I tre nylige Fossil Fredag-artikler (Bechly 2023b, 2024a, 2024b) og en podcast (Bechly 2024c) introduserte jeg (Bechly) en videre ny utfordring for neo-darwinismen, som jeg kalte det kollapsende treproblemet. Dette problemet er utbredt i alle grupper av organismer, og jeg demonstrerte det med eksemplene på fylogeniene til arachnid arthropoder, insektivore pattedyr og moderne (neoaviske) fugler. Den generelle kjernen i argumentet er følgende: Når du ser på de forskjellige fylogenetiske trærne publisert av den mest moderne forskningen på en bestemt gruppe (la oss bare si artiklene de siste ti årene), basert på de mest sofistikerte metodene og de mest komplette dataene, og deretter beregner et såkalt konsensustre som fellesnevner, har disse trærne en tendens til å kollapse til en uavklart busk (polytomi). Dette er til og med innrømmet i hovedstrøms vitenskapelige artikler (f.eks. Sharma et al. 2014). Dette er selvfølgelig hovedsakelig på grunn av de allestedsnærværende motstridende tretopologiene produsert av forskjellige datasett. Dette motsier totalt den nødvendige prediksjonen fra darwinismen om at forskjellige datalinjer skal konvergere til ett ekte livets tre, men det bekrefter greit "plen-synet" fra Darwin -kritikere. Den eneste topologiske strukturen som overlever denne prosedyren er den pre-darwinistiske liné-klassifiseringen, dermed phyla, klasser, ordener og familier vil fremdeles bli bekreftet, men all inter-ordinal og inter-familie-gruppering postulert av darwinisme og "felles avstamming med modifikasjon 'fordamper'. Hovedsakelig skjøt moderne fylogenetikk tilbake mot felles avstamning, eller i det minste svekker de et av de viktigste argumentene som er fremkjempet av darwinistiske aktivister som Richard Dawkins, til fordel for felles avstamning. Dette bør t.o.m. gi darwinister grunn til pause, men i stedet trekker de bare på skuldrene av slike motstridende bevis og forklarer det bort med ad hoc rednings-teknikker som ufullstendig linje-sortering og konvergent evolusjon. Ingen kritikk av det grunnleggende paradigmet er tillatt, ut fra smålige bevis.
I tre nylige Fossil Fredag-artikler (Bechly 2023b, 2024a, 2024b) og en podcast (Bechly 2024c) introduserte jeg (Bechly) en videre ny utfordring for neo-darwinismen, som jeg kalte det kollapsende treproblemet. Dette problemet er utbredt i alle grupper av organismer, og jeg demonstrerte det med eksemplene på fylogeniene til arachnid arthropoder, insektivore pattedyr og moderne (neoaviske) fugler. Den generelle kjernen i argumentet er følgende: Når du ser på de forskjellige fylogenetiske trærne publisert av den mest moderne forskningen på en bestemt gruppe (la oss bare si artiklene de siste ti årene), basert på de mest sofistikerte metodene og de mest komplette dataene, og deretter beregner et såkalt konsensustre som fellesnevner, har disse trærne en tendens til å kollapse til en uavklart busk (polytomi). Dette er til og med innrømmet i hovedstrøms vitenskapelige artikler (f.eks. Sharma et al. 2014). Dette er selvfølgelig hovedsakelig på grunn av de allestedsnærværende motstridende tretopologiene produsert av forskjellige datasett. Dette motsier totalt den nødvendige prediksjonen fra darwinismen om at forskjellige datalinjer skal konvergere til ett ekte livets tre, men det bekrefter greit "plen-synet" fra Darwin -kritikere. Den eneste topologiske strukturen som overlever denne prosedyren er den pre-darwinistiske liné-klassifiseringen, dermed phyla, klasser, ordener og familier vil fremdeles bli bekreftet, men all inter-ordinal og inter-familie-gruppering postulert av darwinisme og "felles avstamming med modifikasjon 'fordamper'. Hovedsakelig skjøt moderne fylogenetikk tilbake mot felles avstamning, eller i det minste svekker de et av de viktigste argumentene som er fremkjempet av darwinistiske aktivister som Richard Dawkins, til fordel for felles avstamning. Dette bør t.o.m. gi darwinister grunn til pause, men i stedet trekker de bare på skuldrene av slike motstridende bevis og forklarer det bort med ad hoc rednings-teknikker som ufullstendig linje-sortering og konvergent evolusjon. Ingen kritikk av det grunnleggende paradigmet er tillatt, ut fra smålige bevis.
Bilde 12. 16 ulike plasseringer for ett og samme objekt -aktuelt empirisk resultat
Fra Darwin tilbake til Linné
 Som en av konsekvensene til det kollapsende treproblemet, foreslår jeg å forlate evolusjons-klassifiseringer og gå tilbake til en pre-darwinistisk liné-klassifisering basert på gruppering i henhold til generell simillitet (fenetikk), og grupper definert av deres unike kombinasjon av diagnostiske tegn, uten noen evolusjonære antagelser. En slik retur til typologisk essensialisme i biosystematikk ville også unngå sirkulære resonnement, som faktisk var et avgjørende argument fra fenetikerne på 1970 -tallet og mønsterkladistene på 1980 -tallet. Dessverre motsto ikke disse to alternative skolene i biosystematikk hovedstrømspresset -mot kladistiske klassifiseringer basert på rekonstruerte evolusjonsforhold. Av hensyn til god vitenskapelig praksis, vil en klassifisering basert på likhetsmønstre måtte komme først, og alle tolkninger skulle først komme senere og være basert på disse primære dataene. Evolusjonister kan selvfølgelig tolke slike likhetsbaserte klassifiseringer når det gjelder felles avstamning med modifisering, og designteoretikere kan alternativt, eller i tillegg, tolke likheter som felles design. Jeg tar ikke til orde for en designbasert klassifisering i stedet for en evolusjonsbasert kladistisk klassifisering, men en teorifri klassifisering basert på observasjonenes empirisk bevis på maksimal generell likhet.
Som en av konsekvensene til det kollapsende treproblemet, foreslår jeg å forlate evolusjons-klassifiseringer og gå tilbake til en pre-darwinistisk liné-klassifisering basert på gruppering i henhold til generell simillitet (fenetikk), og grupper definert av deres unike kombinasjon av diagnostiske tegn, uten noen evolusjonære antagelser. En slik retur til typologisk essensialisme i biosystematikk ville også unngå sirkulære resonnement, som faktisk var et avgjørende argument fra fenetikerne på 1970 -tallet og mønsterkladistene på 1980 -tallet. Dessverre motsto ikke disse to alternative skolene i biosystematikk hovedstrømspresset -mot kladistiske klassifiseringer basert på rekonstruerte evolusjonsforhold. Av hensyn til god vitenskapelig praksis, vil en klassifisering basert på likhetsmønstre måtte komme først, og alle tolkninger skulle først komme senere og være basert på disse primære dataene. Evolusjonister kan selvfølgelig tolke slike likhetsbaserte klassifiseringer når det gjelder felles avstamning med modifisering, og designteoretikere kan alternativt, eller i tillegg, tolke likheter som felles design. Jeg tar ikke til orde for en designbasert klassifisering i stedet for en evolusjonsbasert kladistisk klassifisering, men en teorifri klassifisering basert på observasjonenes empirisk bevis på maksimal generell likhet.
Bilde 13. Carl von Liné -kom med en bestående inndeling
Konklusjoner
 Jeg (Bechly) foreslår at ventetidsproblemet, artsparutfordringen og det kollapsende treproblemet, sammen med de mange andre dyptgripende utfordringene til neo-darwinismen, som de allestedsnærværende konfliktene mellom molekylære klokke-datinger og fossilprotokollen, de voldsomme diskontinuitetene i fossilregisteret (se Bechly 2024D), det kombinatoriske søke-rom-problemet for nye proteinfolder og ikke-reduserbar kompleksitet i molekylære maskiner, har nådd en terskel av motstridende bevis som krever en paradigmendring i biovitenskap og en oppvåkning av teleologi.. Uansett hvilket nytt paradigme som vil erstatte neo-darwinisme, må fordelen av en slik ny tilnærming estimeres ikke bare basert på deres empiriske støtte, årsakssammenheng, generell forklaringskraft, sannsynlighet og parsimonier, men også basert på deres evne til å løse nevnte problemer.
Jeg (Bechly) foreslår at ventetidsproblemet, artsparutfordringen og det kollapsende treproblemet, sammen med de mange andre dyptgripende utfordringene til neo-darwinismen, som de allestedsnærværende konfliktene mellom molekylære klokke-datinger og fossilprotokollen, de voldsomme diskontinuitetene i fossilregisteret (se Bechly 2024D), det kombinatoriske søke-rom-problemet for nye proteinfolder og ikke-reduserbar kompleksitet i molekylære maskiner, har nådd en terskel av motstridende bevis som krever en paradigmendring i biovitenskap og en oppvåkning av teleologi.. Uansett hvilket nytt paradigme som vil erstatte neo-darwinisme, må fordelen av en slik ny tilnærming estimeres ikke bare basert på deres empiriske støtte, årsakssammenheng, generell forklaringskraft, sannsynlighet og parsimonier, men også basert på deres evne til å løse nevnte problemer.
Dette innebærer på ingen måte noen fornektelse av godt etablert vitenskap som geologisk datering og felles avstamming med modifisering, men innebærer en nødvendighet for et nytt syn på mekanismene som fører til biologiske nyheter, som ikke viker unna muligheten for målrettede prosesser. I stedet for å la materialistisk verdenssyn-skjevhet bestemme hvilke forklaringer som er tillatt, og hvilke som blir betraktet som tabu, så skal det bare være de vitenskapelige empiriske dataene i stedet for dogmatiske verdensbilder, som bestemmer blant konkurrerende hypoteser i et søk etter beste forklaring, etter bevisene -uansett hvor det fører.
Bilde 14. Relevant sammenligning
For Litteratur se slutten av originalartikkelen -lenke.
Oversettelse, via google oversetter, og bilder ved Asbjørn E. Lund